 Home
Home Leitfaden
Leitfaden Stoff
Stoff Forum
Forum Links
Links Impressum
ImpressumÜbersicht Teil 3
Die Glykolyse (hier am Beispiel der Leberzelle)
Hier sind wirklich alle aufgeführten Fakten von herausragender Wichtigkeit, da alles auf irgeneiner Weise schon mal abgefragt wurde!
D. h. nicht nur Enzyme und Substrate, sondern auch alle Coenzyme und vor allem wo und auf welcher Höhe bestimmte Substrate aus der Glykolyse
heraus geholt werden können bzw. in welche anderen Stoffwechsel sie eingespeist werden können!
Also: aufmerksam weiterlesen.
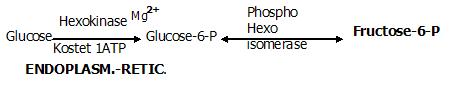
Nach Aufnahme der Glucose in die Zelle wird sie zunächst am C6-Atom im ENDOPLASMATISCHEN RETIKULUM phosphoryliert. Enzym: Hexokinase- Wirkung auf
alle Hexosen mit sehr hoher Aktivität (kleiner Km-Wert) Glukokinase - v. a. Enzym der Leber (und Pankreas), wo es spezifisch auf die Glukose wirkt-
es besitzt eine 100x geringere Aktivität als die Hexokinase(hoher Km-Wert- d.h. zur Aktivität kann es nur dann kommen, wenn das Glukoseangebot entsprechend hoch ist!),
Seine Synthese und somit auch sein Aktivität wird durch Insulin reguliert.
Coenzym: Magnesium
Da die Glukokinasereaktion von Insulin abhängig ist, besteht hier die Möglichkeit die Menge der umgestzten Glukose zu beeinflussen.
Ab einer bestimmten Konzentration an Glukose-6-P stellt die Hexokinase seine Aktivität ein. (Die hier stattfindende Hemmung ist eine allosterische Hemmung = das Enzym ändert
seine Konormation und die Aktivität ist eingestellt)
Dadurch steigt die Glukose-Konzentration in der Leberzelle an. Dies stellt allerdings kein tatsächliches Problem dar, da diese steigende Konzentration und natürlich
auch das vorhandene Insulin, nun die Synthese und Aktivität der Glukokinase steigern.
Mit anderen Worten: Hier besteht eine Regulationsmöglichkeit der Glykolyse, die Erste!
(Weitere Regulationsmöglichkeiten s. u.)
Diese PHOSPHORYLIERUNG hat den einfachen Grund, die Glucose in der Zelle zu halten, da sie nun aufgrund der Ladung die das gebundene Phosphat trägt
die Membran nichtmehr permeieren kann.
Cave: Dieser Schritt wird von einigen Autoren (und auch vom IMPP) nicht zur Glykolyse gerechnet!!!
Außerdem ist er irreversibel (bis auf in der Gluconeogense).
Eine Frage mit der die Studenten gerne auf Glatteis geführt werden:
Welche der Aussagen trifft zu?
a) Die Phosphorylierung der Glucose findet im Zytosol statt
b) Diese Phosphorylierung stellt den ersten Schritt der Glykolyse dar
c) Die Glukokinase ist das als erstes aktiv werdende Enzym
d) Die Glukokinase wird in der ß-Zelle des Pankreas und in den Leberzellen durch Insulin induziert.
Dies ist eine heiß diskutierte Frage, sowohl von den Herren des IMPP als auch von den Lokal-Profs:
(mit der Maus über den schwarzen Balken fahren)
Die eigentliche 1. (reversible) Reaktion
besteht in der Isomerisierung zu Fructose-6-P.
Enzym: Phosphohexose-Isomerase (Glucose-6-P-Isomerase)
Hemmung des Enzyms durch: Kohlenhydratmangel, Glucose-6-P
Eine Aldose (Glucose-6-P) wird zur Ketose (Fructose-6-P)
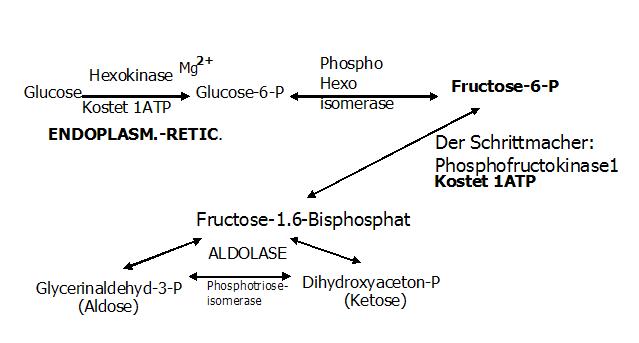
2. Reaktion (exergon + irreversibel)
Phosphorylierung (Knüpfung einer Esterbindung) am C1-Atom des Fructose-6-P zu Fructose-1,6-bisphosphat.
Enzym: Phosphofruktokinase 1 (PFK 1)
Hemmung des Enzyms durch: ATP, Citrat
Verlust eines ATPà ADP+P
Dieser Schritt ist ein sehr wichtiger (weil Schrittmacherreaktion) und wird mit all seinen Zusammenhängen sehr gerne abgefragt!!
Exkurs Schrittmacherreaktion
Diese Reaktion wird deshalb so bezeichnet, weil andere Stoffwechselprodukte auf dieser Höhe in der Lage sind, die Glykolyse zu verlangsamen/ hemmen bzw. zu beschleunigen/ aktivieren. Da die Glykolyse bei der Energiebereitstellung helfen soll, macht es durchaus Sinn sie zu beschleunigen/ aktivieren wenn der ADP-AMP-Spiegel erhöht bzw. die ATP-Citrat-Spiegel erniedrigt sind! Andersrum wird eine Verlangsamung/ Hemmung erreicht, wenn der ADP-AMP-Spiegel erniedrigt ist bzw. ATP-Citrat-Spiegel erhöht.Außerdem sollte man sich klar machen, dass diese Art der Regulierung sich auf die Beeinflussung des Enzyms bezieht! Entweder es wird allosterisch gehemmt oder aktiviert.
Was ist falsch?
a) Die PFK1 (Phosphofruktokinase) ist ein Schlüsselenzym der Glykolyse.
b) Die PFK1 wird durch ATP und Citrat gehemmt.
c) Die PFK1 wird durch Fruktose-2,6-Bisphosphat aktiviert.
d) Die PFK1 wird durch ADP und AMP gehemmt.
Gesuchte Falschaussage: (mit der Maus über den schwarzen Balken fahren)
3. Reaktion (Bildung einer Schiff-Base)
Zwischen Enzym und Substrat kommt es zur Bildung einer Schiff´schen Base. Dadurch kann der 6er-Zucker in zwei 3er-Zucker gespalten werden.
Substrat: 6er-Zucker, Fructose-1,6-bisphosphat
Enzym: Aldolase
Ergebnis: 2 Triosen
- Glycerinaldehyd-3-P (GAP=Aldotriose)
- Dihydroxyaceton-P (DAP=Ketotriose)
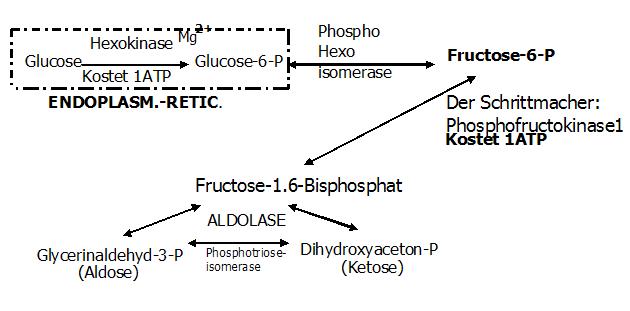
Exkurs Aldolase
Aus Fructose-1,6-bisphosphat entstehen 2 Triosen.
1. GAP
2. DAP
Das katalysierende Enzym ist eine Aldolase,
die Fructose-1,6-bisphosphat-Aldolase.
Dieses besteht aus 4 Untereinheiten. Diese wiederum sind aus den
Isoformen A, B und C zusammengesetzt.
Isoformen A+C haben eine hohe Spezifität für Fructose-1,6-Bisphosphat
Isoform B - als einziges in der Lage Fructose-1-Phosphat zu spalten- zentrale Rolle im Fructosestoffwechsel, Exprimierung in Leber und Niere (postnatal),- bei Defekt
dieser-----hereditäre Fructose-Intoleranz mit den Symptomen Schwitzen und Krämpfe als Zeichen der Hypoglykämie nach Fruktosegabe.
Zwischen der Carbonylgruppe der F-1,6-BP und der E-Aminogruppe der Aldolase, kommt es zur Ausbildung einer SCHIFF BASE,
die zu einer Lockerung der Bindung der Fructose zwischen dem C3- und dem C4-Atom führt, mit der Möglichkeit sie zu spalten!
zu GAP
zu DAP (kann von Muskel-u. Fettgewebe zur TG-Synthese verwendet werden)
Es ist sehr wichtig zu verstehen, dass aus einer Hexose zwei Triosen entstanden sind. Und ab diesem Moment der Trennung bzw. der Bildung dieser Beiden,
logischerweise die Glykolyse auch zweimal durchlaufen werden muß (jede Triose einmal). Und das wiederum bedeutet, dass sich die Energiebeute ab diesem Moment auch verdoppelt!!
4.Reaktion (reversibel)
Diese beiden Triosen stehen miteinander über eine gemeinsame Enolform im Gleichgewicht. Dieses Gleichgewicht liegt weit aif der Seite des DAPs!
Zum weiteren Ablauf der Glykolyse wird jedoch GAP benötigt. Die Einstellung des Gleichgewichtes wird durch die Triosephosphatisomerase beschleunigt,
die eine sehr hohe Umsatzzahl besitzt und so Glycerophosphat, das nur in sehr kleinen Mengen vorliegt, schnell nachliefern kann.
Enzym: Triosephosphatisomerase
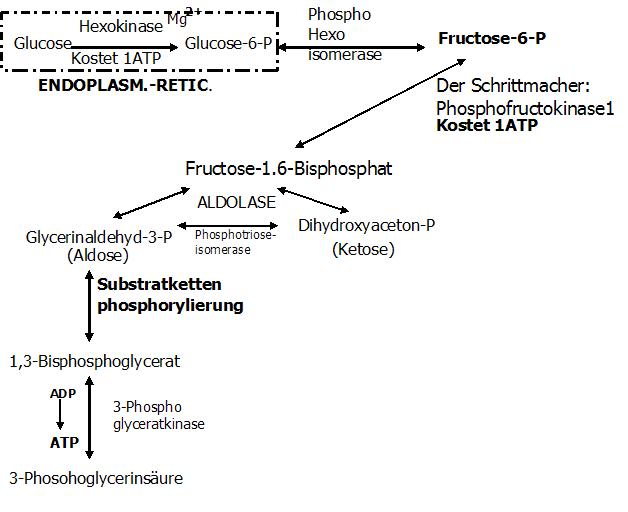
5.Reaktion (Substratkettenphosphorylierung + reversibel)
Das C1-Atom des GAPs wird oxidiert.
Enzym: 3-Phosphoglycerinaldehyd-Dehydrogense (Hemmung des Enzyms durch: NADH2)
Coenzym: NAD
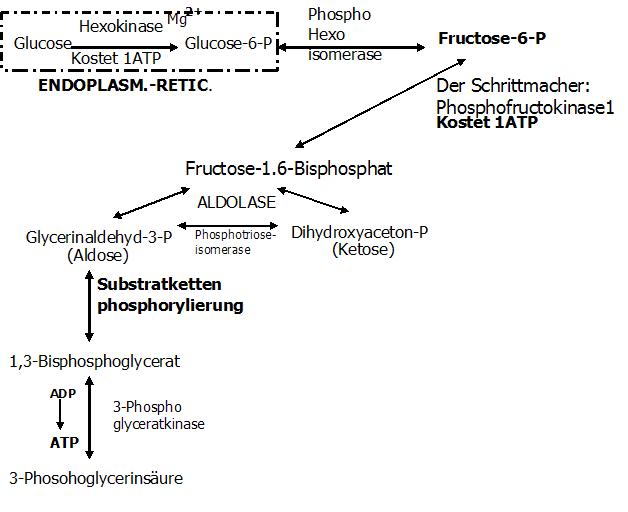
Exkurs Substratkettenphosphorylierung
Durch die Substratkettenphosphorylierung kommt es zur Konservierung der Energie. An die SH-Gruppe des Enzyms wird die Carbonylgruppe (C1-Atom) des GAPs gebunden.
Es entsteht ein Thioester.
Bei dieser Bindung werden 2H frei die aufgefangen werden müssen. Dies übernimmt das Coenzym NAD, welches mit dem Enzym zu Beginn der Reaktion zu einem
Enzym-NAD-Komplex zusammengelagert ist. Es entstehen ein NADH+H und ein Substrat-Enzym-Komplex.
Dieses NADH2 wird in die Atmungskette geschleust, wo es für die Energieausbeute von 3ATPs verantwortlich ist. Gewinn des NADH2àAtmungskette (à Gewinn von 3ATP)
Nun werden Enzym und Substrat voneinander getrennt. Phosphorylytische Spaltung. Das Ergebnis ist das 1,3-Bisphosphoglycerat.
Das heißt dieses Substrat trägt 2 Phosphatgruppen, von denen nun eins energiereich ist!!
6.Reaktion: Gewinn eines ATP (1 ATP pro Triose)
Der Phosphatrest der energiereich am C1-Atom gebunden ist, kann direkt auf ADP übertragen werden.
Enzym: 3-Phosphoglyceratkinase
Ergebnis: 3-Phosphoglycerinsäure
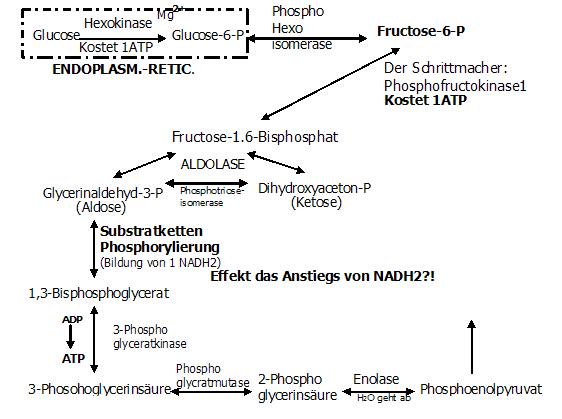
7. Reaktion: Vorbereitung des P um noch mehr Energie bereitzustellen (reversibel)
Da die 3-Phosphoglycerinsäure noch ein anorganisches P besitzt, wäre es natürlich am sinnvollsten,
es dem Stoffwechsel angemessen zuzuführen... nämlich als Energie!!
Also muß dieses P auf die Energieaufnahme vorbereitet bzw. es muß aktiviert. Dies geschieht, indem das P vom C3 auf das C2 Umgelagert wird.
Enzym: Phosphoglyceratmutase
Ergebnis: 2-Phosphoglycerinsäure
Man kann also sagen, daß das Phosphat lediglich als Speichermöglichkeit funktioniert (sowie der Kanister das Benzin speichert).
8. Reaktion: Der Sinn dieser Reaktion liegt darin, das vorbereitete P jetzt mit Energie zu beladen.
Enzym: Enolase (auch Phosphoenolathydratase/ Lyase)
Ergebnis: Phosphoenolpyruvat
Die Energie selber ensteht dadurch, daß die Enolase H2O abspaltet und es somit zu einer Umverteilung der Energie (in Form von Wärme) kommt,
die ja sowieso in diesem Substrat enthalten ist!!!
Diese Energie wird letzendlich in Form der Ester-Bindung, zwischen dem P und der O-Gruppe des Phosphoenolpyruvat gespeichert.
(Also, nicht das Phosphat oder das ATP ist die Energie,
sondern die Verbindung zwischen dem P und dem jeweiligen Substrat stellt die Energie... in Form von Wärme.)
9. Reaktion: ADP ---> ATP (irreversibel)
Phosphoenolpyruvat wird unter ATP-Bildung zu Pyruvat umgewandelt
(Irreversibel ist diese Reaktion, da das Phosphoenolpyruvat ein höheres Phosphoryl-Gruppenübertragungspotential beitzt als das ATP.)
Enzym: Pyruvatkinase
Hemmung des Enzyms durch: ATP, Citrat
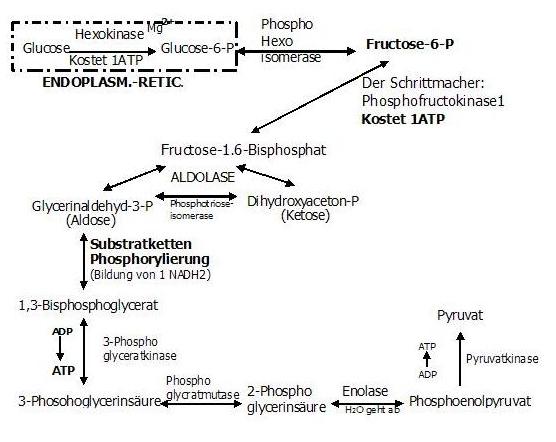
Nun fehlt noch eine letzte Reaktion und wir haben´s geschafft!!
10. Reaktion: ANAEROBE GLYKOLYSE
Dazu stellen wir uns folgende Situation vor:
Unsere Zellen befinden sich in einer ANAEROBEN Stoffwechsellage (z. B. weil wir gerade dabei sind wie der "Wilde Bill" ums Haus zu laufen oder
weil wir uns in glücklichen/ misslichen Lage irgendwelcher emotionaler Empfindungen befinden)
Erinnere: Die Atmungskette-Komplex I-V
HIER SEI AUSDRÜCKLICH AUF DIE HERRAUSRAGENDEN EINFLUß DES 02 AUF DIE ATMUNGSKETTE HINGEWIESEN, DIE OHNE DEN SAUERSTOFF LOGISCHERWEISE NICHT ABLAUFEN KANN!!
D. h. dass diese Reaktion nur dann abläuft wenn die Zelle nicht genügend Sauerstoff zur Verfügung hat!!
Herrscht also O2-Mangel (s. o.), so kann das aus Reaktion 5 entstehende NADH2, seine beiden Hs nicht an die Atmungskette weiter geben!
Somit ergibt sich folgendes Problem:
Das sich nun anstauende NADH2 würde die 5. Reaktion am Ablauf behindern
(durch Hemmung des Enzyms: 3-Phospho-Glycerinaldehy-Dehydrogenase)!
Dies ist der Grund, weshalb sich die Evolution die "Anaerobe Glykolyse" ausgedacht hat.
Denn durch diese 10. Reaktion kann das NADH2 "entsorgt" werden.
Enzym: Lactatdehydrogenase (LDH)
Ergebnis: Lactat
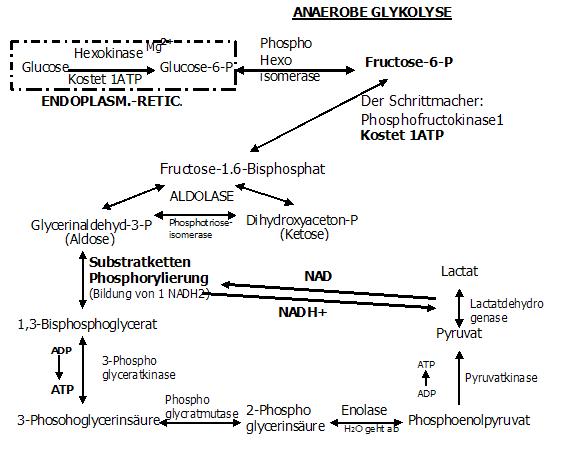
Um die Verwirrung komplett zu machen, gehen wir jetzt auf die Regulation der Glykolyse ein.
Dazu folgende Fragen:
Wann bzw. warum ist es sinnvoll für die Glykolyse abzulaufen?
- v. a. wenn genug Glucose vorhanden ist
- wenn in der Zelle ein Mangel an ATP und auch an Citrat vorherrscht bzw. die ADP-Konzentration ansteigt
Wie lange sollte sie ablaufen?
- bis der Energiemangel aufgehoben ist oder die Zelle auf dem besten Weg dahin ist will sagen, dass die Vorstufen der Energie oder sie selbst in ihrer Konzentration ansteigen (z. B. Pyruvatà Acetyl CoA, Citrat, ATP); diese Substrate sind in der Lage, die Schlüsselenzyme der Glykolyse zu hemmen. Wann sollte sie nicht mehr ablaufen ja sogar daran gehindert werden!?
- wenn der Energiemangel vollends gedeckt ist bzw. viel ATP und somit wenig ADP
- wenn die Energiespeicher (Glykogen in Leber (90%) und Muskel (10%) aufgefüllt sind
Welche besonderen Faktoren sollten Sinnvollerweise Einfluß auf diese Regulation haben?
Nun auch dieses ist ein ganz wichtiges Thema, welches sehr gern von dem IMPP zu unserem Leid herangezogen wird.
Wie wir gerade gelernt haben, verringert sich der Umsatz der Glykolyse oder kommt auch ganz zum erliegen, wenn die Konzentration insbesondere des ATP groß genug ist.
Das liegt daran, dass das ATP einen Hemmeffekt auf die PFK1 hat. Der Sinn dieser Hemmung liegt darin, nicht zu viel ATP zu bilden (so wie es die Menge an der
vorhandenen Glucose durchaus erlauben würde). Denn ATP hat eine leider zu kurze Halbwertszeit t1/2 und ATP kann auch nicht gespeichert werden. Dementsprechend wäre
alles ATP das gebildet aber nicht verbraucht dem unmittelbaren Zerfall geweiht. Also wird die Aktivität der Glykolyse gebremst bis gehemmt. (Und nicht vergessen,
dass wir uns noch immer in der Leber befinden!)
Frage:
Ist die letzendliche Bildung von ATP (Glukose à Pyruvatà Acetyl-CoA à Citratcyklus à Energie bzw. ATP) die einzige Möglichkeit um die Glukose im Stoffwechsel zu verwerten??
Wohl mit Sicherheit nicht (wenn denn schon mal so penetrant danach gefragt wird)!!
Wir haben im Kapitel der Übersicht schon gelernt, dass das Endprodukt der aeroben Glykolyse (Pyruvat) in die PDH eingeschleust wird, um als Acetyl-CoA den Weg
weiter zu beschreiten! Einerseits im Citratcyklus und andererseits kann es noch zu FETTSÄURE weiter verstoffwechselt werden!
Also wäre es ja gar nicht so schlimm wenn die Glykolyse weiter ihren regulären Weg durchsetzen würde, da das entstehende Pyruvatà Acetyl-CoA ja noch
die genialen Ausweichmöglichkeit hat und zur Fettsäure werden kann!
Fettsäure: alternativer und genialer Energiespeicher!!
Da dieser Speicher unbegrenzt gefüllt werden kann (das sieht man manchen echt gut an!) ist es gar nicht so schlimm wenn die Glykolyse weiter ablaufen würde,
im Gegenteil, es wäre sogar sinnvoll! Denn somit würde die Glucose, die ja in ihrer Konzentration immer weiter ansteigt, sinnvoll
verwertet werden (als Fettsäuren-Energiespeicher) Also musste sich die Evolution etwas ausdenken, um diesen ATP-Hemmeffekt aufheben zu können.
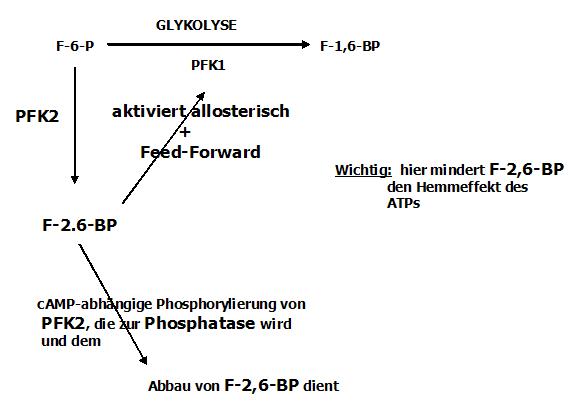
Die Hemmung des Hemmeffektes des ATP auf das Schlüsselenzym PFK1
Ist also der Umsatz groß, so ist u. a. die Synthese des Substrates Fructose-6-P gesteigert. Diese erhöhte Konzentration führt nun dazu, dass ein weiteres
Enzym in der Leber (das unabhängig von der Glykolyse ist) aktiviert wird: die Phosphofruktokinase 2
Diese phosphoryliert die Fructose-6-P in Fructose-2-6-bisphosphat.
Dieses Substrat hat nun 2 wichtige Aufgaben
- Die Wiederaktivierung des Schlüsselenzyms PFK1
- Verminderung des ATP-Hemmeffektes auf die PFK1
Entschuldigt bitte, Übersicht Teil 4 ist in Arbeit